
期刊:CELL
影响因子:66.85
技术服务:scRNA-seq
导语
尽管以T细胞为中心的免疫疗法已经取得了无可争议的临床成功,但数量有限的获得持久响应的患者迫切需要补充的治疗策略。NK细胞,作为肿瘤微环境中一类重要的组成部分参与肿瘤控制的多个过程,如直接的杀伤细胞和分泌促炎因子等。利用NK细胞进行癌症治疗已提出了大量的策略,其特点是有希望的特性,包括其安全性和有效性。特别值得注意的是,嵌合抗原受体(CAR)- NK细胞治疗淋巴瘤、骨髓瘤和白血病取得了显著的临床成功。然而,基于NK细胞的治疗在实体瘤中受到阻碍,部分原因是对肿瘤浸润性NK细胞的了解不完全,特别是它们对肿瘤的浸润、表型异质性和TME内的失调。
人体中的NK细胞可被划分为2种主要的类,CD56dimCD16hi和CD56brightCD16lo,主要基于CD56 (NCAM1) 和 CD16 (FCGR3A)的表达水平。CD56dimCD16hi NK细胞群主要通过分泌穿孔素和颗粒酶介导靶细胞的杀伤,而CD56brightCD16lo NK细胞群表现出免疫调节和细胞因子产生能力。近期,单细胞RNA测序技术促进了肿瘤浸润免疫细胞异质性的表征,为阐明肿瘤浸润NK细胞亚群的图谱提供了很大机会。例如,我们鉴定了CD160+HSPA1A+肝常驻细胞亚型在肝细胞癌中的特异性富集,其他人则报道了出现在黑色素瘤中浸润的特异的NK细胞亚群具有区域性差异。同时,人类NK细胞在在健康组织和血液中的分布和功能也得到了研究。然而,对于肿瘤浸润的NK细胞,大多数单细胞RNA测序的规模是有限的,并且它们在恶性疾病中获得的异质性表型的程度仍不清楚。
与CD8+ T细胞相比,NK细胞作为细胞毒性活性的替代来源,以低突变负荷和主要组织相容性复合体(MHC) I类异常表达来对抗肿瘤细胞。尽管CD8+ T细胞的功能失调状态以细胞毒性降低和多种抑制受体的高表达为特征,但NK细胞功能失调尚未得到详细研究。此前,肝癌中有NK细胞功能低下的报道,但尚未分析其免疫调节机制,其他癌症类型是否也存在这种现象尚不清楚。此外,尽管TIGIT和TIM3在肿瘤浸润性NK细胞中的抑制作用已经明确,但其他免疫检查点如PD-1和CTLA4是否在这些细胞中发挥同样的作用甚至表达仍然存在争议。此外,NK细胞对TME的免疫抑制因子敏感,这可能导致它们的功能失调的表型,但不同的调节过程如何影响不同癌症类型的TME中NK细胞的功能和丰度尚不清楚。总之,这些促使我们对NK细胞进行深入研究,以阐明它们在泛癌症水平上的异质性和功能障碍。
在这里,我们收集了广泛的已发表和新生成的scRNA-seq数据,构建了一个全面的肿瘤浸润人类NK细胞图谱,并探索了NK细胞在不同癌症类型和组织中的异质性。我们揭示了肿瘤浸润NK细胞状态的转变,并强调了可能导致NK细胞功能障碍的TME成分。这些数据将为促进对主要癌症类型中NK细胞的整体特性的理解提供丰富的资源,并为基于NK细胞的免疫治疗的发展提供有价值的见解。
技术服务
scRNA-seq
研究结果
1. 在单细胞分辨率水平下构建人类泛癌NK细胞图谱
为肿瘤浸润NK细胞构建一个综合的泛癌单细胞转录组图谱,我们首先从我们新生成的数据集中收集了47名诊断为8种癌症类型之一的患者的scRNA-seq数据和70个其他已发表的数据集。这些数据覆盖了24种癌症类型,包括来自716个病人的1223个样本,包括肿瘤、临近非肿瘤组织、外周血、和其他组织如淋巴结,以及60名健康对照样本(图1A-B)。经过严格的质控和计算门控(CD3CD56+/KLRF1+)及非监督聚类结合的方法,本研究中我们获得了160011个高质量的NK细胞,包含11963个新生成的NK细胞。值得注意的是,考虑到肿瘤组织内NK细胞在CD45+细胞中相对较少,对它们的异质性了解的较少,因此,构建如此大规模的NK细胞图谱是必不可少且有挑战性的。与先前的来自人血液和脾脏的7000个NK细胞构建的NK细胞单细胞转录组图谱相比,我们的规模扩大了20倍,代表了大多数主要的癌症类型和多种组织。
为了无偏的定义NK细胞的泛癌群体结构,我们整合了数据集之间最小批次效应的单细胞RNA数据,并进行了2轮无监督聚类。第一轮分析涉及区分两种特征明确的主要细胞类型,CD56brightCD16lo 和 CD56dimCD16hi,主要基于高表达的典型细胞标记NCAM1和FCGR3A,分别对应于先前报道的“NK_1”和“NK_2”群体。CD56brightCD16lo区可以进一步细分为5个子集,而CD56dimCD16hi区在第二轮聚类中被识别出9个子集(图1C-D)。我们没有观察到两种主要大群在不同癌症类型或不同亚群中KLRF1的表达有显著差异. 值得注意的是,之前报道的ILC细胞标记基因几乎不在这些NK细胞亚群中表达,表明了我们数据的纯度。这些亚群的特点是在每个主要群体中都有不同的特征基因的高表达。正如预期的那样,先前描述的亚群很容易在我们的图谱中被识别出来,例如CD56brightCD16lo c5-CREM亚群伴随着CREM高表达。伴随着KLRC2 (NKG2C)的高表达的CD56dimCD16hi c8-KLRC2 NK 细胞亚群被视为适应性的NK细胞。我们的图谱还发现了几个被低估的NK细胞亚群,它们具有独特的转录表型。例如,CD56dimCD16hi c5-MKI67以MKI67和STMN1等增殖标志物的高表达为特征,CD56dimCD16hi c6-DNAJB1特异性表达与应激反应相关的基因。我们的图谱还在几乎所有癌症类型中捕获了低比例的CD56brightCD16hi NK细胞,同时表达高水平的NCAM1和FCGR3A。CD56brightCD16hi NK细胞表现出介于CD56brightCD16lo和CD56dimCD16hi NK细胞之间的中间特征,可能代表了类似于小鼠CD27+CD11b+ NK细胞的发育中间体,这也被认为是小鼠的短暂成熟阶段,但scRNA-seq几乎无法检测到。接下来,我们检查了所有亚群的组织分布,观察到不同的组织富集模式,表明我们的综合分析可以保留不同组织的异质性 (图1E-F)。为了进一步证实聚类的稳定性,我们将血液、肿瘤和邻近非肿瘤组织中的NK细胞分别重新聚类,结果显示高度一致。最后,利用可以测量细胞簇纯度的全局未移位熵比(ROGUE)指数,表明所有这些群体在各种癌症类型中都是稳健的。
NK细胞亚群参与了不同的发育阶段,其共同谱系特异性基因的表达揭示了这一点。肿瘤中富集的CD56bright CD16lo c4-IL-7R和血液中富集的c2-IL-7R-RGS1lo细胞,高表达NK细胞前体标志物,都是在发育早期绘制的。相比之下,其他未成熟的CD56brightCD16lo亚群表现出前体标志物的逐渐减少和CD160表达的大量增加。更成熟的CD56dimCD16hi亚群,特别是c6-DNAJB1和c7- NR4A3,除了FCGR3A和B3GAT1 (CD57)外,还表现出杀伤免疫球蛋白样受体(KIR)家族的上调。不同发育状态的NK细胞亚群在肿瘤中同时存在,表明NK细胞向肿瘤的迁移可能与NK细胞成熟解耦有关。
我们进一步检查了基因表达特征,以破译不同群体之间的功能差异(图1G)。CD56dimCD16hi NK细胞高表达细胞毒效应基因,包括穿孔素(PRF1)和除GZMK外的大多数颗粒酶(GZMB、GZMA和GZMH),而GZMK只在CD56brightCD16lo NK细胞中表达。CD56brightCD16lo NK细胞表达多种细胞因子基因,如IL18。我们还观察到CD56brightCD16lo c2和c4同时表达il -18及其受体IL18R1,这意味着il -18依赖性自分泌途径在这些亚群中可能起着至关重要的作用。引人注目的是,CD56dimCD16hi NK细胞亚群也可以表现出某些细胞因子的特异性表达,但与CD56brightCD16lo亚群具有不同的模式。特别是,CD56dim CD16hi亚群c4-NFKBIA在所有NK细胞亚群中表现出相对较高的炎症评分,主要表达CCL3、CCL4和CCL4L2,表明它们有能力招募其他免疫细胞,如T细胞。最近的研究将1型常规树突状细胞(cDC1s)募集到肿瘤与NK细胞联系起来,通过NK细胞分泌XCL1、XCL2和CCL5来实现。我们的研究鉴定出了通过细胞类型特异性互补策略参与cDC1募集的多种NK细胞亚群。CD56brightCD16lo c2和c4表达初级水平的XCL1和XCL2,而CD56brightCD16lo (c1-GZMH和c3-CCL3)和CD56dimCD16hi (c1-IL-32和c8-KLRC2) NK细胞优先表达另一个cDC1趋化基因CCL5。此外,我们描述了激活和抑制受体的概况,观察到NK细胞亚群的明显变化。例如,KLRC1在CD56brightCD16lo NK细胞中的表达水平高于CD56dimCD16hi NK细胞,尽管不是特异性的,LAG3和TIGIT在c8-KLRC2中高表达。值得注意的是,与其他组织富集的CD56dimCD16hi亚群(c4-NFKBIA, c5-MKI67和c7-NR4A3)相比,c6-DNAJB1 NK细胞表现出最高的应激评分和最弱的细胞毒性,表明它们的转录和功能表型存在显著差异(图1G)。综上所述,我们以细粒度的亚群分辨率提供了NK细胞的详细转录组谱。而不是分析NK细胞作为一个单一的群体,我们剖析NK细胞的亚群特异性分子特性,并揭示其未被重视的异质性。
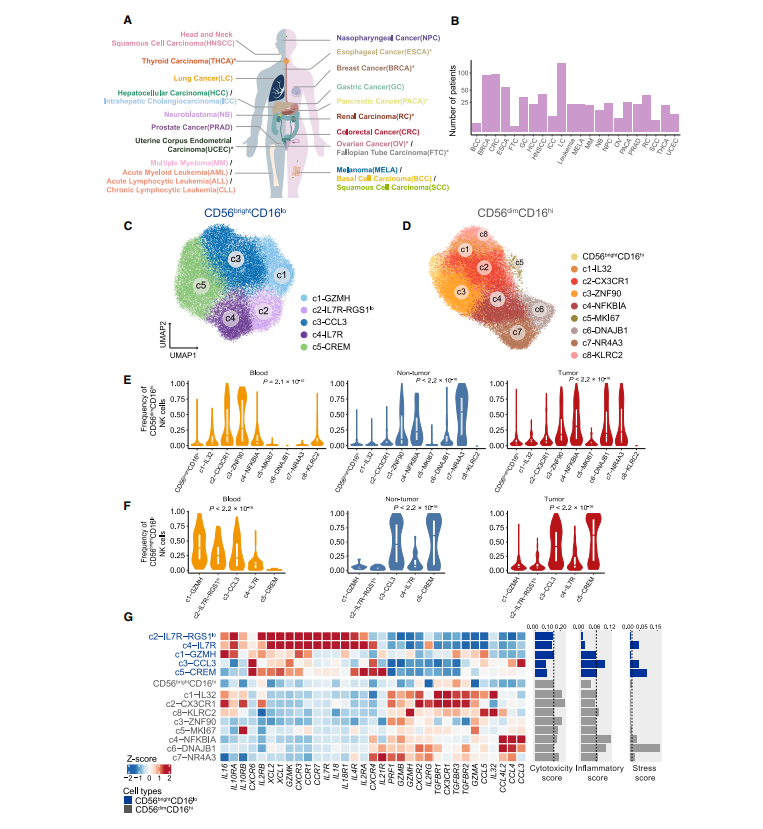
图1
2. 不同肿瘤类型NK细胞的组织异质性
接下来,我们评估了肿瘤浸润NK细胞群在不同癌症类型中的偏好,观察到明显的差异(Figure 2A)。例如,未成熟的CD56brightCD16lo NK细胞在鼻咽癌和基底细胞癌中占主导地位,而成熟的CD56dimCD16hi NK细胞在肾癌和肺癌中占主导地位,这与先前的报道一致。其他的,如结直肠癌和肝癌,没有明显的倾向(图2A)。为了检验上述趋势是否可以用器官结构来解释,我们分析了邻近非肿瘤组织中主要NK细胞类型的内在组成及其在肿瘤中的相应变化。在某些类型的癌症中,如结直肠癌,肿瘤组织的NK细胞组成与邻近的非肿瘤组织相似。在肺癌、肾癌等肿瘤类型中,尽管肿瘤与邻近非肿瘤组织中主要NK细胞类型保持不变,但CD56dimCD16hi NK细胞的比例明显降低(图2B)。有趣的是,乳腺癌和食管癌肿瘤主要的NK细胞群与它们的邻近非肿瘤组织相比是相反的(图2C)。这些观察结果表明,内部器官特性和恶性肿瘤相关因素对形成NK细胞群的成分有复合的影响。
我们进一步从子集的角度探索NK细胞的癌症类型特异性,并进行无监督聚类,根据NK细胞亚群比例对所分析的癌症类型进行分层。NK细胞亚群如c2-CX3CR1在胰腺癌、乳腺癌和黑色素瘤中表现出强烈的偏好。在某些亚型的癌症类型中观察到高变异性。例如c4-NFKBIA和c7-NR4A3从头颈部鳞状细胞癌和甲状腺癌到食管癌和鼻咽癌的中位数频率显著降低(图2D)。尽管观察到明显的差异,但包括子宫内膜癌、基底细胞癌和食管癌在内的癌症类型聚集在一起,均具有丰富的CD56brightCD16hi c5-CREM和较少的c4-IL-7R NK细胞。特别值得注意的是,具有上述低饱和度阶段的罕见CD56brightCD16hi NK细胞亚群在黑色素瘤和白血病中大量存在,特别是在急性髓系白血病(AML)亚型中(图2D)。NK细胞的这种低饱和度阶段与AML患者的总生存期和无复发生存期的降低有关。与其他NK细胞群相比,这些CD56brightCD16hi细胞在其极低的激活和抑制受体评分方面表现出独特的表型和功能变化(图2E)。目前基于NK细胞的治疗侧重于增强NK细胞的激活和寿命,但通常忽略了癌症类型之间的异质性和TME对NK细胞细胞毒性功能的抑制作用,这些都应该在未来的治疗策略中加以考虑。
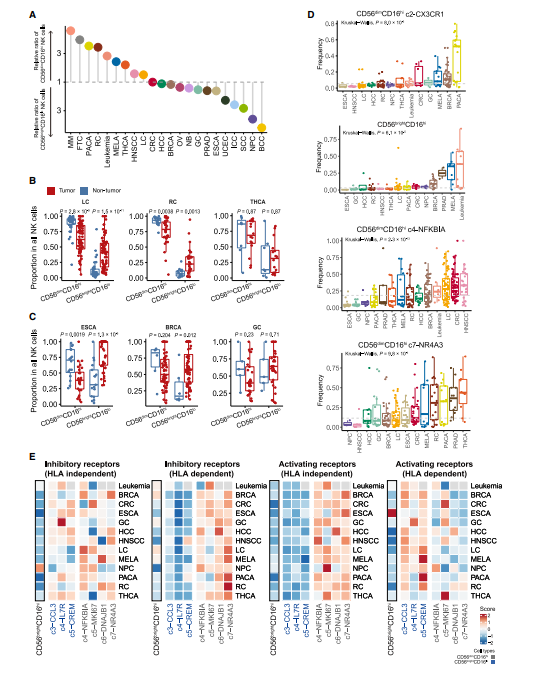
图2
3. RGS1是组织浸润NK细胞的标志
如上所述,NK细胞成分从血液到组织都发生了显著变化。同样,在组织浸润NK细胞和它们的血液对应物之间检测到广泛的转录变化(图3A-B)。先前的研究已经确定了组织驻留NK细胞的几种标记物,如CD69、CD103、CXCR6和CD49a。然而,我们发现ITGA1 (CD49a)、ITGAE (CD103)和CXCR6在单细胞转录组水平很少被检测到,CD69在NK细胞中广泛表达,包括血液来源的(图3E)。此外,有报道显示,这些标记物在特定组织的NK细胞群中优先表达。这些促使我们以无偏的方式从泛癌的角度发现强大的NK细胞驻留标记。
我们选择了血液和组织之间的差异表达基因,并进一步评估了它们的敏感性和特异性,以区分NK细胞的组织来源。因此,RGS1 (G蛋白信号1的调节因子)被明确识别,它只在肿瘤和邻近非肿瘤组织的NK细胞中表达,而在血液中几乎检测不到(图3C-D)。此外,RGS1的表达与KLF2、SELL等迁移信号相反(图3E)。与上述常规组织驻留标记相比,RGS1具有更高的敏感性和特异性。接下来,我们直接比较了RGS1与ITGAE、CD69及其联合在血液中的表达模式,发现RGS1单独在血液中的检出率最低,而RGS1/CD69联合可以进一步提高组织中的阳性检出率(图3F)。此外,RGS1/CD69联合在区分血液和非血液NK细胞方面比单独使用具有更高的曲线下面积(AUC)值。值得注意的是,RGS1在分析的患者和癌症类型中广泛表达(图3G-H)。
综上所述,这些特征暗示了RGS1单独或与CD69结合的潜在作用,在转录组水平上是组织浸润NK细胞的优良标记物。我们推测RGS1的表达可能减弱G蛋白的信号活性,导致NK细胞趋化迁移能力减弱,促进NK细胞驻留。RGS1在NK细胞中的作用机制有待进一步研究。
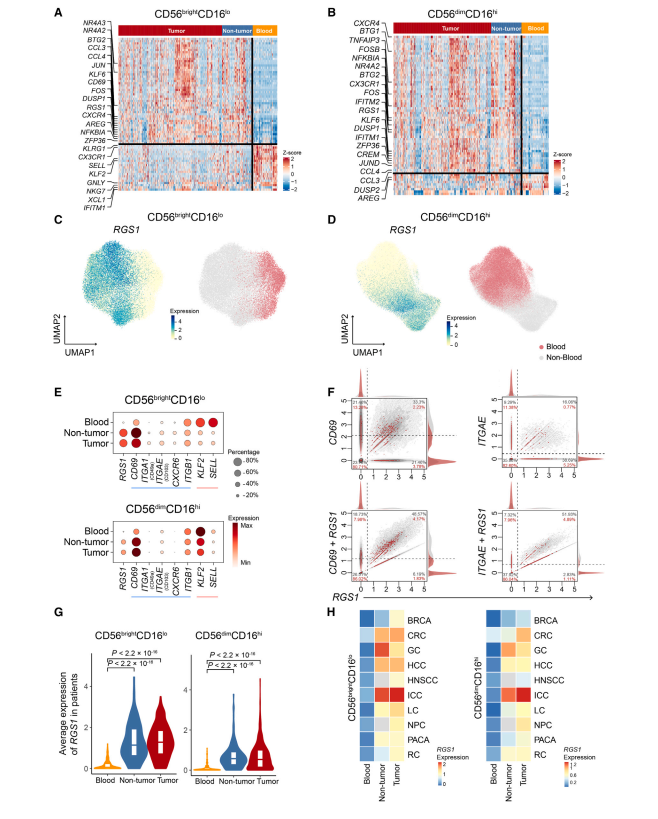
图3
4. 肿瘤相关NK细胞程序及其特征
接下来,我们试图阐明NK细胞在肿瘤中的明确特征。除了上述某些亚群在肿瘤特异性富集外,我们发现,与邻近的非肿瘤组织相比,在肿瘤中,CD56brightCD16lo c3-CCL3 NK细胞的细胞因子产量较低,表现为CCL3和CCL4的表达减少,并且在大多数癌症类型中,c5-CREM NK细胞中XCL1和XCL2的表达减少,暗示它们在肿瘤中的功能转换。然后,我们使用SCENIC识别了肿瘤浸润性CD56brightCD16lo和CD56dim CD16hi NK细胞亚群的激活调控子。重要的是,肿瘤富集的c6-DNAJB1亚群表现出更高的转录因子如KLF6和EGR3的表达,这些转录因子与细胞毒性功能的抑制有关。
利用RNA速率,我们解码了NK细胞的转录动力学,在CD56brightCD16lo和CD56dimCD16hi NK细胞中观察到从血液富集的亚群到肿瘤浸润群的明确定向流动(图4A)。相应的,RGS1的表达沿流速方向升高。我们发现CD56dimCD16hi c6-DNAJB1 NK细胞位于速率末端,从而推断为终末状态(图4A)。值得注意的是,来自邻近非肿瘤组织的CD56dimCD16hi NK细胞主要观察到在UMPA图上富含c7-NR4A3细胞; 相比之下,肿瘤来源的CD56dimCD16hi NK细胞在富含c6-DNAJB1细胞的UMAP图上占优势地位(图4B)。与此一致的是,肿瘤富集的c6- DNAJB1 NK细胞的标记物,如DNAJB1和HSPA1A在肿瘤浸润的CD56dimCD16hi NK细胞群中高度表达(图4C)。此外,我们观察到c6-DNAJB1和c7-NR4A3 NK细胞中线粒体基因的表达水平相似,表明c6-DNAJB1 NK细胞的应激表型与细胞质量无关。由于c6-DNAJB1细胞在肿瘤中特异性富集,我们将这一群体称为肿瘤相关NK (TaNK)细胞。
多重免疫荧光染色进一步证实了肿瘤中TaNK细胞的存在(图4D)。我们还利用流式细胞术验证了TaNK细胞(CD56dimCD16hi HSP40+)在体内肿瘤的富集。在1例肝内胆管癌和6例HCC样本中,我们发现了TaNK细胞,它们在肿瘤浸润性CD56dimCD16hi NK细胞中的比例高于对应的邻近肝组织,这与我们的scRNA-seq数据一致(图4E)。
然后,我们使用拟时序推断分析来研究CD56dimCD16hi NK细胞的动态,发现TaNK细胞在CD56dimCD16hi NK细胞的推断拟时间内越来越多地出现,并且在终末期富集,与RNA速率分析的结果一致。为了检验TaNK细胞的新特征,我们将基因表达谱与拟时间拟合。有趣的是,CD56dimCD16hi NK细胞在过渡过程中表现出细胞毒性降低,抑制受体和应激基因表达升高(图4F)。
值得注意的是,在所有肿瘤浸润的CD56dimCD16hi NK细胞亚群中,末端TaNK细胞具有最低的细胞毒性和最高的应激评分。相比之下,相应的c7- NR4A3在邻近的非肿瘤组织中富集,具有高度的细胞毒性(图 4G)。通过对几种癌症类型进行多重免疫荧光染色,我们观察到TaNK细胞表现出较低水平的GZMB (图4H)。肝癌患者流式细胞术分析进一步证实,与CD56dimCD16hi HSP40+ NK细胞相比,TaNK细胞肿瘤部位细胞毒性颗粒(颗粒酶B和穿孔素)表达较低,抑制受体CD158a (KIR2DL1)和CD158e (KIR3DL1)表达较高(图4I -J)。这些结果表明,TaNK细胞可能与功能失调状态有关。此外,我们还观察到NR4A核受体家族随着拟时间顺序的增加而呈现的动态差异趋势(图4F)。c7-NR4A3 NK细胞高表达NR4A2和NR4A3,而TaNK细胞高表达NR4A1。有趣的是,NR4A1已被确定为T细胞功能障碍的关键介质,并被认为有助于限制实体瘤中CAR - T细胞的功能。总之,我们的数据表明,肿瘤中的TaNK细胞可能最终功能失调,并可能在TME中发挥关键作用。

图4
5. TaNK细胞与不良预后和免疫治疗耐药的关系
由于NK细胞和CD8+ T细胞表现出广泛的表型和功能相似性,我们接下来研究了靶向CD8+ T细胞的免疫检查点阻断(ICB)疗法是否也会影响NK细胞。在肿瘤浸润性NK细胞和CD8+ T细胞中,TaNK细胞和耗竭T细胞(Tex)表现出明显的应激状态(图5A),提示它们参与肿瘤免疫应答。均高度表达一系列抑制受体分子;然而,它们在各种免疫调节基因上具有不同的表达谱。传统的免疫检查点基因如PDCD1和CTLA4在TaNK细胞上几乎不表达(图5A),这意味着它们不是抗PD -1/CTLA-4治疗的直接靶点。因此,在TME和当前的ICB治疗中,TaNK细胞可能与Tex细胞发挥不同的作用。
我们观察到不同癌症类型的TaNK细胞丰度存在显著差异(图5B),肿瘤时期对TaNK细胞比例的影响很小(图5C)。值得注意的是,在癌症基因组图谱(TCGA)数据集中,肿瘤中的高TaNK细胞信号与大多数癌症类型的低生存率相关(图5D-E)。我们进一步应用基于深度学习的模型进行反卷积和细胞组成分析,发现高TaNK细胞频率表明癌症患者预后不良。此外,我们通过分析先前乳腺癌和黑色素瘤的ICB治疗研究中预处理肿瘤的scRNA-seq数据,进一步研究了TaNK细胞是否与ICB治疗应答相关(图5F)。引人注目的是,在两种癌症类型中,无应答患者中观察到的TaNK细胞比例高于应答患者。进一步利用来自各种癌症(包括黑色素瘤、肺癌、转移性尿路上皮癌)的已发表的大量数据,我们验证了无应答患者比应答患者表现出更强的TaNK细胞信号(图5G)。
我们推测,长期浸润可能赋予肿瘤中TaNK细胞的功能状态,导致其对恶性细胞的无效杀伤。TaNK细胞的富集与对抗肿瘤的免疫反应受损以及对当前ICB治疗的低敏感性有关。我们的发现揭示了TaNK细胞在肿瘤中的潜在作用,并为NK细胞免疫疗法的合理设计提供了参考。
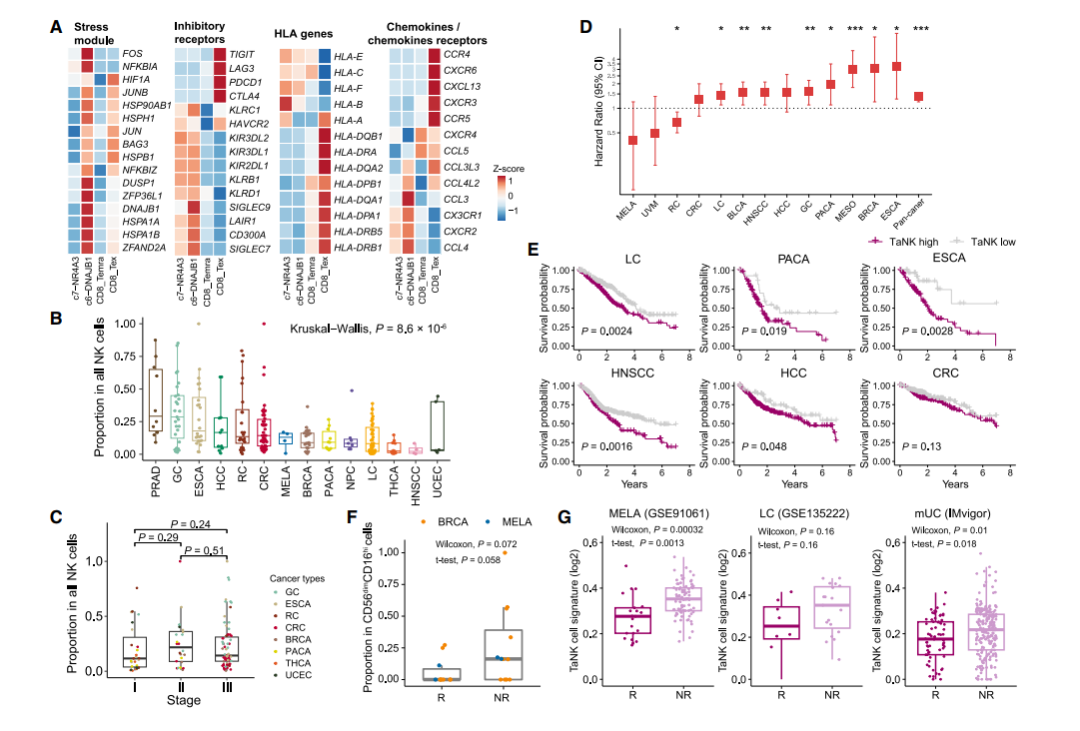
图5
6. TME中塑造肿瘤浸润NK细胞功能的潜在介质
为了深入了解NK细胞在TME中的调控程序,我们利用CellPhoneDB来探测NK细胞和其他CD45+免疫细胞(包括T细胞和髓系细胞)之间潜在的细胞-细胞相互作用。与T细胞相比,除肥大细胞外,大多数髓系细胞类型与CD56dimCD16hi NK亚群表现出强烈的潜在相互作用(图6A)。特别有趣的是,TaNK细胞被预测通过ANXA1调节多种髓系细胞类型,ANXA1是一种在炎症反应中与免疫抑制和诱导巨噬细胞重编程相关的蛋白质(图6B)。这表明功能失调的NK细胞可能具有抑制TME中促炎巨噬细胞的潜力。为了进一步阐明NK细胞来源的ANXA1在巨噬细胞中的作用,我们对肺癌和肝癌的肿瘤样本进行了多重免疫荧光染色,发现了一个ANXA1+ NK细胞群。与远离ANXA1+ NK细胞的巨噬细胞相比,靠近ANXA1+ NK细胞的巨噬细胞表现出较低的活化标志物CD86(图6C-D)和较高的抗炎标志物转化生长因子b (TGF-b)的表达水平(图6E-F)。
值得注意的是,在树突状细胞(DC)亚群中,LAMP3+ DC,最近表征的成熟cDCs(也称为mregDC)显示出与CD56dimCD16hi NK细胞最强的相互作用潜力(图6A)。多重免疫荧光分析显示,LAMP3+ DCs与NK细胞共定位(图6J)。此外,预测它们之间的相互作用通过IL-15-IL-15受体和NECTIN2-TIGIT相互作用轴介导(图6B)。重要的是,LAMP3+ DC在转录组水平上表达免疫群体中最高水平的IL15、PVRL2 (NECTIN2)和PVR(图6G)。流式细胞术也证实了IL-15在LAMP3+ DC中的高表达(图6H)。IL-15已被确定为维持NK细胞寿命的稳态相关细胞因子,并用于NK细胞输注和体外繁殖。相比之下,TIGIT作为抑制受体有助于抑制NK细胞介导的免疫反应。此外,在TCGA数据集中,LAMP3+ DC的丰度与CD56dimCD16hi NK细胞相关(图6I)。接下来,我们研究了LAMP3+ DC在肿瘤中的具体调控过程,发现肿瘤浸润的LAMP3+DC与邻近非肿瘤组织相比,IL15的表达较低(图S6K),这表明LAMP3+ DC可能对TME中CD56dimCD16hi NK细胞的激活作用受损。事实上,物理上接近LAMP3+ DC的NK细胞表达颗粒酶B的水平较低(图6J-K)。总之,我们的分析表明,在TME中,LAMP3+ DC对CD56dimCD16hi NK细胞的异常调节。
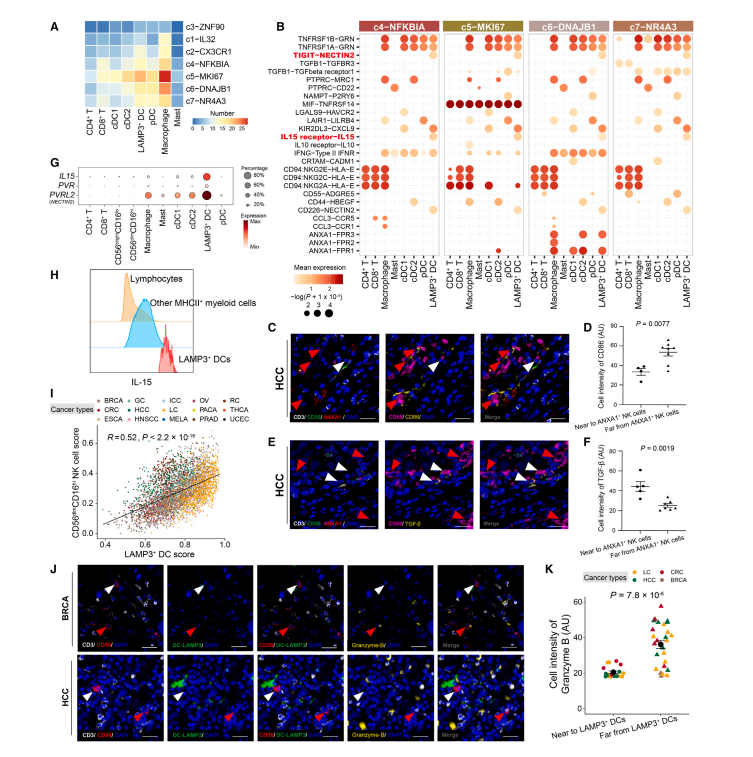
图6
7. 外周血NK细胞亚群的不同转录组模式
NK细胞在血液中的淋巴细胞室中占相当大的比例,但它们在肿瘤诱导的外周免疫系统中的作用相对不明确。我们的图谱包含来自35名健康供者的血液来源NK细胞的9个scRNA数据集,使我们能够探测肿瘤患者外周血中NK细胞的特异性改变。
我们首先比较了来自健康供者的循环NK细胞与来自肿瘤患者的循环NK细胞的转录组特征,不同数据集的健康供者的循环NK细胞显示出高度相似性(图7B)。相比之下,对于来自肿瘤患者的循环CD56brightCD16lo细胞,我们观察到与健康供者的转录组存在显著差异;在所分析的所有癌症类型中,循环CD56dimCD16hi细胞的差异甚至更为显著(图7A)。值得注意的是,肿瘤患者在NK细胞亚群中表现出显著的组成变化,这种模式似乎是癌症类型特异性的(图7C)。例如,在结直肠癌、头颈部鳞状细胞癌、肾癌和HCC中,循环CD56brightCD16lo c3-CCL3 NK细胞的比例增加,但在其他被分析的癌症类型中没有增加。
接下来,我们将重点放在CD56dimCD16hi c8-KLRC2适应性NK细胞上,该细胞在某些癌症类型如结直肠癌和胃癌中富集(图7D)。适应性NK细胞被认为是CAR NK细胞的一个有吸引力的来源,因为它们具有增强细胞因子反应的效应特性和对免疫抑制效应的内在抗性。特别有趣的是,根据我们的数据,与其他循环NK细胞相比,这些细胞特异性表达MHC II类基因(图7E)。我们还检查了这些NK细胞在肿瘤患者中的功能变化,发现与健康供体相比,患者来源的适应性NK细胞的功能基因和MHC II类基因的表达明显更高(图7F, 7H),这意味着它们在肿瘤患者中处于高度激活状态。我们通过流式细胞术进一步证实,与健康供者相比,HCC患者循环NK细胞中MHC II类分子的高表达(图7G)。因此,在患者源性适应性NK细胞中上调的基因参与了免疫效应过程的正调控等途径(图7I)。综上所述,我们的分析表明,循环NK细胞参与了肿瘤进展过程中外周免疫环境的系统性变化。

图7
结论
NK细胞在对抗肿瘤进展的先天免疫应答中起着不可或缺的作用。为了描述他们在肿瘤微环境中的表型和功能多样性,我们对716名癌症患者的NK细胞做了综合的单细胞RNA测序分析,涵盖了24种癌症类型。我们在肿瘤类型特异的种类中观察到NK细胞组成的异质性。值得注意的是,我们鉴定出了一组肿瘤相关的NK细胞,他们在肿瘤中富集,显示出抗肿瘤功能受损,并与不良预后及免疫治疗耐受有关。特定的髓系细胞亚群,特别是LAMP3+DC细胞,可能介导NK细胞抗肿瘤免疫的调节。总之,我们的综合分析增强了目前从泛癌症角度对NK细胞的理解,阐明了NK细胞群体结构以及肿瘤诱导的局部和全身NK细胞反应的见解。为了方便广泛的研究社区使用我们的数据,已经开发了一个交互式门户网站(http://pan-nk.cancer-pku.cn/)来分析和可视化我们的单细胞数据。我们设想我们的大规模数据可以进一步促进NK细胞免疫疗法在更多癌症患者中的应用。
参考文献:
Tang F, Li J, Qi L, Liu D, Bo Y, Qin S, Miao Y, Yu K, Hou W, Li J, Peng J, Tian Z, Zhu L, Peng H, Wang D, Zhang Z. 2023. A pan-cancer single-cell panorama of human natural killer cells. Cell, 186(19):4235-4251. doi: 10.1016/j.cell.2023.07.034.